Ryosuke Omori et Jianhong Wu développent un algorithme inductif pour étudier les fréquences de nucléotides spécifiques au site à l'aide d'un modèle multi-souche susceptible-infectieux-éliminé (SIR) pour mieux comprendre l'épistémologie des maladies infectieuses, évolution des agents pathogènes, et la dynamique des populations. Crédit :Wikimedia Commons.
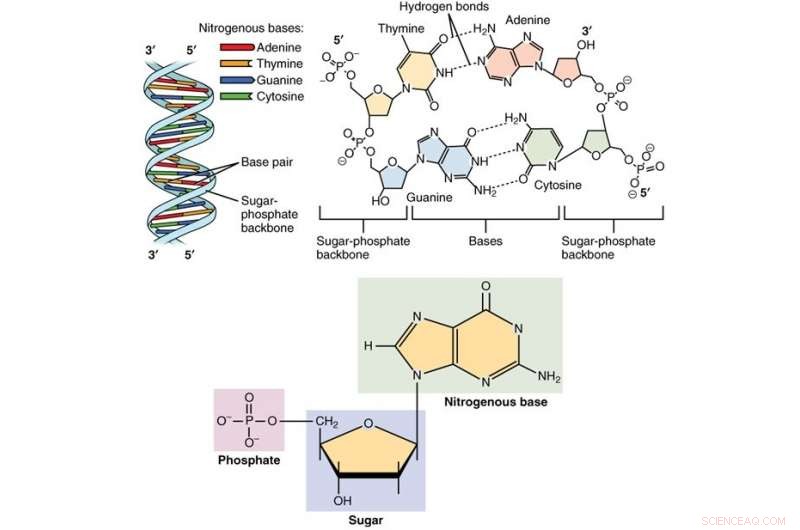
Méthodes de séquençage des acides nucléiques, qui déterminent l'ordre des nucléotides dans les fragments d'ADN, progressent rapidement. Ces processus produisent de grandes quantités de données de séquence, dont certaines sont dynamiques, qui aident les chercheurs à comprendre comment et pourquoi les organismes fonctionnent comme ils le font. Le séquençage profite également aux études épidémiologiques, telles que l'identification, diagnostic, et le traitement des maladies génétiques et/ou contagieuses. Les technologies de séquençage avancées révèlent des informations précieuses sur l'évolution temporelle des séquences d'agents pathogènes. Parce que les chercheurs peuvent estimer comment une mutation se comporte sous la pression de la sélection naturelle, ils sont ainsi capables de prédire l'impact de chaque mutation - en termes de survie et de propagation - sur la fitness du pathogène en question. Ces prédictions donnent un aperçu de l'épistémologie des maladies infectieuses, évolution des agents pathogènes, et la dynamique des populations.
Dans un article publié plus tôt ce mois-ci dans le Revue SIAM de Mathématiques Appliquées , Ryosuke Omori et Jianhong Wu développent un algorithme inductif pour étudier les fréquences de nucléotides spécifiques à un site à l'aide d'un modèle multi-souche susceptible-infectieux-éliminé (SIR). Un modèle SIR est un modèle compartimental simple qui place chaque individu d'une population à un moment donné dans l'une des trois catégories susmentionnées pour calculer le nombre théorique de personnes touchées par une maladie infectieuse. Les auteurs utilisent leur algorithme pour calculer le D de Tajima, un test statistique populaire qui mesure la sélection naturelle sur un site spécifique en analysant les différences dans un échantillon de séquences d'une population. En situation non endémique, Le D de Tajima peut changer avec le temps. L'étude de l'évolution temporelle du D de Tajima au cours d'une épidémie permet aux chercheurs d'estimer les mutations pertinentes pour la fitness des agents pathogènes. Omori et Wu visent à comprendre l'impact de la dynamique de la maladie sur le D de Tajima, conduisant ainsi à une meilleure compréhension de la pathogénicité d'une mutation, gravité, et la spécificité de l'hôte.
Le signe du D de Tajima est déterminé à la fois par la sélection naturelle et la dynamique des populations. "Le D de Tajima est égal à 0 si l'évolution est neutre - pas de sélection naturelle et une taille de population constante, " dit Omori. " Une valeur non nulle du D de Tajima suggère une sélection naturelle et/ou un changement dans la taille de la population. Si aucune sélection naturelle ne peut être supposée, Le D de Tajima est fonction de la taille de la population. D'où, il peut être utilisé pour estimer les changements de séries chronologiques de la taille de la population, c'est à dire., comment évolue l'épidémie."
Équations différentielles, qui modélisent les taux de variation du nombre d'individus dans chaque compartiment du modèle, peut décrire la dynamique des populations. Dans ce cas, la dynamique des populations d'hôtes infectés par la souche portant une séquence donnée est modélisée par un ensemble d'équations différentielles pour cette séquence, qui incluent des termes décrivant le taux de mutation d'une séquence à une autre. Lors de la configuration de leur modèle SIR multi-souches, Omori et Wu supposent que la dynamique de population de l'agent pathogène est proportionnelle à la dynamique de la maladie. c'est à dire., le nombre d'agents pathogènes est proportionnel au nombre d'hôtes infectés. Cette hypothèse permet à la valeur du D de Tajima de changer.
En génétique des populations, les chercheurs pensent que le signe du D de Tajima est affecté par la dynamique de la population. Cependant, les auteurs montrent que dans le cas d'un modèle déterministe SIR, Le D de Tajima est indépendant de la dynamique de la maladie (en particulier, indépendamment des paramètres du taux de transmission de la maladie et du taux de guérison de la maladie). Ils observent également que si le D de Tajima est souvent négatif au début d'une épidémie, il devient souvent positif avec le temps. "Le signe négatif n'implique pas une expansion de la population infectée dans un modèle déterministe, " a déclaré Omori. " Nous avons également constaté que la dépendance du D de Tajima sur la dynamique de transmission de la maladie peut être attribuée à la stochasticité de la dynamique de transmission au niveau de la population. Cette dépendance est différente de l'hypothèse existante susmentionnée sur la relation entre la dynamique de la population et le signe du D de Tajima."
Finalement, Omori et Wu prouvent que le D de Tajima dans un modèle SIR déterministe est complètement déterminé par le taux de mutation et la taille de l'échantillon, et que l'évolution temporelle de la diversité génétique d'un agent pathogène de maladie infectieuse est entièrement déterminée par le taux de mutation. "Ce travail a révélé une certaine dépendance du D de Tajima sur le nombre de reproduction de base (dynamique de transmission de la maladie) (R
Compte tenu de la demande d'outils d'analyse de la dynamique évolutive et de la maladie, l'observation que le D de Tajima dépend de la stochasticité de la dynamique est utile pour estimer les paramètres épidémiologiques. Par exemple, si des séquences d'agents pathogènes sont échantillonnées à partir d'un petit foyer dans une population hôte limitée, alors le D de Tajima dépend à la fois du taux de mutation et du R