Parce que le parasite du paludisme Plasmodium ne peut pas prévoir quand il peut être transmis d'un moustique à un hôte mammifère, il utilise des protéines de liaison poly(A) spécialisées pour emballer et protéger son matériel génétique à utiliser après la transmission. Crédit : Centers for Disease Control and Prevention
Le parasite qui cause le paludisme n'en a pas, mais deux, des protéines spécialisées qui protègent ses ARN messagers (matériel génétique qui code pour les protéines) jusqu'à ce que le parasite s'installe dans un nouveau moustique ou un hôte humain. Une nouvelle étude menée par des chercheurs de Penn State décrit les deux protéines et révèle un rôle supplémentaire que l'on peut jouer pour faciliter les interactions basées sur l'ARN entre le parasite, son moustique vecteur, et son hôte humain. L'étude paraît le 10 janvier 2018, dans la revue mSphère .
« Comprendre le parasite du paludisme et la façon dont il interagit avec son hôte peut fournir des informations qui pourraient aider à prévenir la propagation de cette maladie souvent mortelle, " a déclaré Scott Lindner, professeur adjoint de biochimie et de biologie moléculaire à Penn State et auteur principal de l'étude. « Le parasite du paludisme a un cycle de vie complexe qui comprend des phases chez le moustique vecteur, le foie humain, et dans le sang humain. De plus, le parasite n'a aucune idée quand il va être transmis d'un moustique à un hôte humain et vice-versa, il doit donc toujours être prêt à être transmis. Il s'y prépare en fabriquant et en emballant les ARNm dont il aura éventuellement besoin pour fabriquer des protéines à l'intérieur de son nouvel hôte ou d'un nouveau moustique. »
Au cours de ce processus, appelé refoulement traductionnel, des protéines spéciales se lient aux ARNm et les empêchent d'être traduits en protéines. Une protéine impliquée se lie à la queue poly(A) de l'ARNm - une chaîne répétée de molécules d'As ou d'adénosine ajoutée à l'extrémité de la plupart des brins d'ARNm. Cela aide à former un complexe de protéines et d'ARN qui est réduit au silence mais prêt à agir après la transmission du parasite à l'hôte. La plupart des organismes unicellulaires ont un type de cette protéine de liaison poly(A), tandis que les organismes multicellulaires en ont deux. Dans cette étude, les chercheurs caractérisent deux types de protéines de liaison poly(A) dans le parasite unicellulaire Plasmodium, tous deux contribuent à la régulation translationnelle.
« Nous savions grâce aux travaux antérieurs de notre laboratoire que Plasmodium avait un type de protéine de liaison poly(A) qui fonctionne en dehors du noyau de la cellule, " a déclaré Allen Minns, technicien de recherche à Penn State et premier auteur de l'article. "Cette protéine se lie et protège la queue poly(A) à une extrémité d'un brin d'ARNm. Dans cette étude, nous avons utilisé des approches biochimiques pour caractériser davantage cette protéine, et a découvert qu'il avait également un travail spécialisé dans la réception d'ARNm. Il forme des chaînes sans la présence d'ARN, ce qui permet potentiellement à de grands assemblages de la protéine de protéger rapidement toute la longueur de la queue poly(A)."
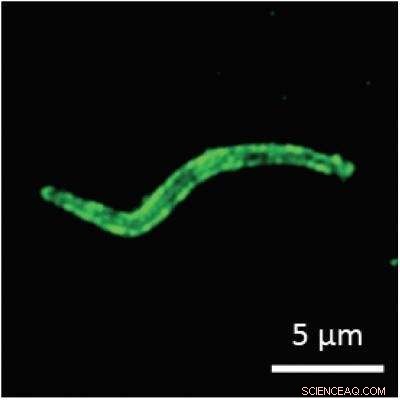
Le sporozoïte du paludisme, étiqueté avec un colorant fluorescent dans cette image, contient la forme non nucléaire d'une protéine de liaison poly(A) à sa surface. Le rôle inattendu de cette protéine sur la forme infectieuse du parasite du paludisme n'est pas encore clair mais peut fournir une opportunité pour le parasite d'interagir avec son moustique vecteur ou son hôte humain via l'ARN. Crédit :État de Pennsylvanie
Les chercheurs ont également identifié et caractérisé un deuxième type de protéine de liaison poly(A) qui fonctionne à l'intérieur du noyau du parasite pendant les étapes sanguines de son cycle de vie. Dans les organismes multicellulaires, cette deuxième protéine de liaison poly(A) effectue généralement un contrôle de qualité avant que l'ARNm ne quitte le noyau, confirmant que l'ARNm est construit correctement. Ces protéines de contrôle qualité transmettent ensuite le brin d'ARNm à d'autres protéines en dehors du noyau, qui dirigent l'ARNm à traduire ou à encapsuler pour une utilisation ultérieure par répression traductionnelle.
En plus d'un rôle important dans la régulation de la traduction à l'intérieur de la cellule, les chercheurs ont également découvert que la protéine de liaison poly(A) non nucléaire peut jouer un rôle surprenant en dehors de la cellule.
"Lorsque le parasite prend la forme d'un sporozoïte chez le moustique, en fait, nous ne voyons pas la grande majorité de la protéine de liaison poly(A) non nucléaire à l'intérieur de la cellule où nous nous attendions à ce qu'elle se trouve - où elle interagirait avec les ARNm produits par le parasite, " dit Lindner. " Au lieu de cela, la protéine s'accumule à la surface du sporozoïte et est éliminée lorsque le parasite se déplace. Nous ne voyons pas cela se produire à d'autres stades de la vie du parasite, et c'est maintenant la troisième protéine de liaison à l'ARN trouvée à la surface du sporozoïte. Le parasite met ces protéines de liaison à l'ARN à sa surface pour une raison; la nouvelle et passionnante question est pourquoi. »
Les chercheurs pensent que les protéines de liaison poly(A) à la surface du sporozoïte permettent au parasite d'interagir avec l'ARN provenant de sources extérieures au parasite et pourraient ainsi fournir une opportunité au parasite d'interagir avec le moustique ou l'hôte via leur ARN.
"Cette étude suggère que l'interaction du parasite avec l'ARN extérieur est probablement beaucoup plus répandue que nous ne le pensions, " a déclaré Lindner. " Il est possible que ce type d'interaction puisse éventuellement fournir une nouvelle cible pour les stratégies d'intervention, mais la première étape consiste à comprendre pourquoi le parasite du paludisme possède ces protéines de liaison poly(A) à la surface du sporozoïte."