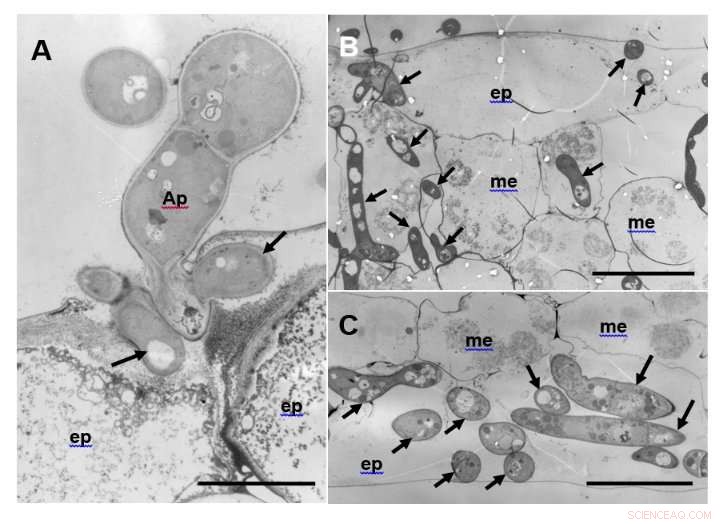
Figure 1 :Images de microscopie électronique à transmission (MET) de feuilles sensibles de R108-1 3 jours après l'inoculation avec M. pinodes. (A) Hyphes d'infection (flèches) qui ressemblent à des vésicules d'infection formées dans la paroi cellulaire de la cellule épidermique adaxiale (ep). Appressorium (Ap). Barre =5 µm. (B) Epiderme adaxial et cellules du mésophylle (me) envahies par les hyphes. Les organites des cellules hôtes ont été dégradés. Barre =20 µm. (C) Hyphes étendus dans les cellules épidermiques abaxiales. Barre =20 µm. Crédit :Université d'Okayama
Ascochyta (Mycosphaerella) pourriture du pois, causée par Mycosphaerella pinodes, est l'une des maladies les plus importantes des légumineuses à grains dans le monde. Malgré l'impact économique et les nombreuses études sur cette maladie, on sait peu de choses sur les caractéristiques cytologiques au cours de l'infection, surtout dans les interactions résistantes. Une des raisons est le manque de cultivars de pois résistants, ainsi que les ressources disponibles dans la collection de matériel génétique Pisum avec une forte résistance à cette maladie.
Kazuhiro Toyoda et ses collègues de l'Université d'Okayama ont examiné l'histologie et l'ultrastructure des événements infectieux précoces et du développement fongique, y compris pénétration par appressoria, croissance végétative des hyphes d'infection et réponses de l'hôte, utilisant un pathosystème modèle récemment développé impliquant Medicago truncatula et la souche OMP-1 de M. pinodes (Toyoda et al., 2013).
Sur l'écotype sensible R108-1, les pycnospores ont germé et se sont développées à la surface de l'épiderme, puis formait une appressoria et pénétrait la cuticule. Sous la cuticule, la cheville d'infection s'est étendue en un hyphe qui s'est développé dans la paroi externe de l'épiderme. Ensuite, les hyphes ont pénétré dans les cellules du mésophylle et ont proliféré vigoureusement, formant éventuellement des fructifications asexuées (pycnides) (Fig. 1).
En revanche, la pénétration réussie et la croissance subséquente des hyphes d'infection étaient considérablement restreintes dans l'écotype Calife (Fig. 2). De façon intéressante, les hyphes aberrants tels que les hyphes intrahyphes et les hyphes morts, grâce à une défense locale provoquée par le champignon, étaient abondants dans Calife mais pas dans R108-1. Détecté par sa réaction avec le chlorure de cérium (CeCl3) pour générer des perhydroxydes de cérium denses aux électrons dans les micrographies électroniques à transmission, le peroxyde d'hydrogène (H2O2) s'est accumulé dans les cellules épidermiques et mésophylles de Calife provoquées par des pycnospores de M. pinodes. Cette localisation intracellulaire a été confirmée par spectroscopie à rayons X à dispersion d'énergie (EDX) (Fig. 3). Ces observations indiquent donc que la réaction d'explosion oxydative conduisant à la génération d'espèces réactives de l'oxygène est associée à une réponse de défense locale de l'hôte chez Calife, puisqu'aucune accumulation claire de H2O2 n'était détectable dans le R108-1 sensible.
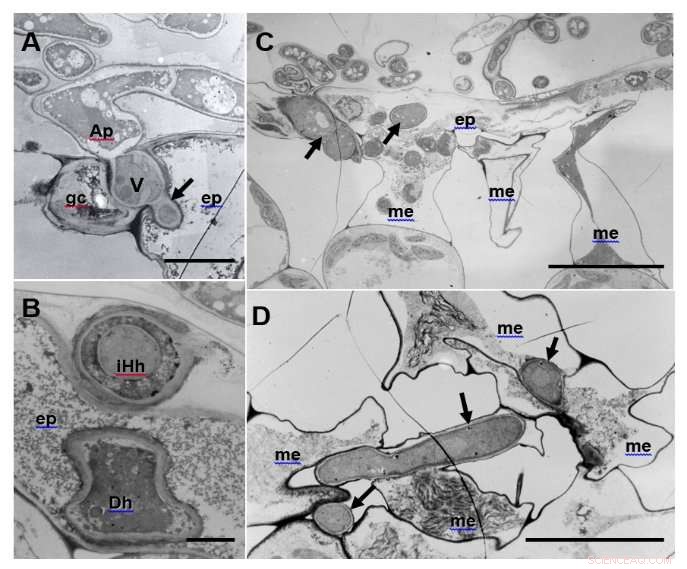
Figure 2 :Images MET de feuilles de calife résistantes 3 jours après l'inoculation avec M. pinodes. (A) La vésicule d'infection (V) a émergé de la pointe d'un appressorium (Ap) aux parois cellulaires entre la cellule de garde (gc) et la cellule épidermique (ep), et les hyphes d'infection se sont étendus dans la cellule épidermique (ep). Barre =5 µm. (B) Hyphes aberrantes dans la cellule épidermique ; hyphes intrahyphes (iHh) et hyphes morts (Dh). Barre =2 µm. (C) L'épiderme envahi par les hyphes et les cellules du mésophylle adjacentes avait rétréci. Barre =20 µm. (D) Les cellules du mésophylle en contact avec les hyphes ont rétréci. Barre =10 µm. Crédit :Université d'Okayama
Les chercheurs concluent que les aberrations structurelles sont probablement des mécanismes courants des champignons pour se protéger d'un environnement hostile chez un hôte résistant via une enceinte par un autre hyphe. Les différences structurelles entre les interactions sensibles et résistantes ainsi que les réponses de l'hôte aideront à mieux comprendre la pathogenèse du champignon sur le pois, fournissant ainsi des informations sur la sélection des cultivars résistants de pois.
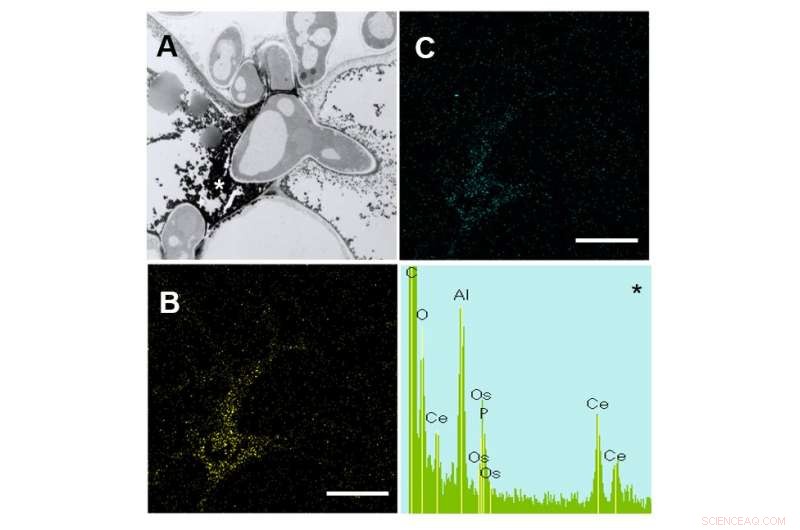
Figure 3 :Images MET de feuilles de calife résistantes 3 jours après l'inoculation avec M. pinodes. (A) La vésicule d'infection (V) a émergé de la pointe d'un appressorium (Ap) aux parois cellulaires entre la cellule de garde (gc) et la cellule épidermique (ep), et les hyphes d'infection se sont étendus dans la cellule épidermique (ep). Barre =5 µm. (B) Hyphes aberrantes dans la cellule épidermique ; hyphes intrahyphes (iHh) et hyphes morts (Dh). Barre =2 µm. (C) L'épiderme envahi par les hyphes et les cellules du mésophylle adjacentes avait rétréci. Barre =20 µm. (D) Les cellules du mésophylle en contact avec les hyphes ont rétréci. Barre =10 µm. Crédit :Université d'Okayama














