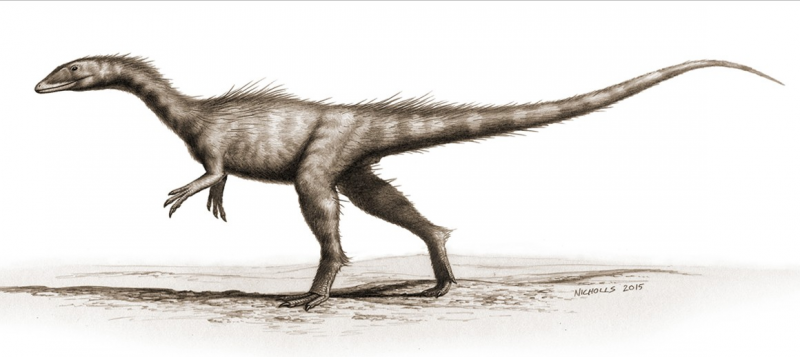
Dracoraptor hanigani (théropode) restauré en tant que prédateur et charognard vivant sur le rivage. Oeuvre de Bob Nicholls (paleocreations.com). Crédit :Muséum d'histoire naturelle, Londres. Crédit :Avancées scientifiques, doi:10.1126/sciadv.aaw4486
L'évolution de l'endothermie (thermorégulation par voie métabolique) représente une transition majeure dans l'histoire des vertébrés. Cependant, le processus d'évolution de l'endothermie et sa chronologie chez les oiseaux et les mammifères restent controversés. Dans un nouveau rapport sur Avancées scientifiques , Enrico L. Rezende et une équipe de chercheurs du Centre d'écologie appliquée et de durabilité, et l'Institut des sciences de l'environnement et de l'évolution au Chili, combiné un modèle de transfert de chaleur avec des données sur la taille du corps des théropodes. Les chercheurs ont ensuite reconstitué l'évolution des taux métaboliques le long de la lignée des tiges des oiseaux. Les résultats suggèrent qu'une réduction de la taille constitue le chemin de moindre résistance pour l'évolution de l'endothermie, maximisant l'expansion thermique de la niche, tout en réduisant les coûts des besoins énergétiques élevés.
Par conséquent, les chercheurs émettent l'hypothèse que le métabolisme aurait augmenté avec la miniaturisation au cours de la période du Jurassique inférieur et moyen (il y a environ 180 à 170 millions d'années) pour entraîner un gradient de niveaux métaboliques dans la phylogénie des théropodes. Alors que les théropodes basaux peuvent avoir présenté des taux de métabolisme plus faibles, les lignées non aviaires les plus récentes étaient probablement des thermorégulateurs décents avec un métabolisme amélioré. L'analyse a fourni une séquence provisoire dans le temps des transitions évolutives clés, pour l'émergence de petits, dinosaures endothermiques et volants à plumes.
L'évolution de l'endothermie chez les oiseaux et les mammifères est une transition importante au cours de l'évolution des vertébrés, fournissant un exemple extraordinaire de convergence évolutive entre les groupes, essentiel à leur distribution géographique étendue et à leur succès écologique. Bien que plusieurs groupes d'invertébrés et de vertébrés puissent élever leur température au-dessus de la température ambiante, la capacité de maintenir une température corporelle élevée et constante via la production de chaleur endogène au repos est exclusive aux oiseaux et aux mammifères seuls. L'observation expliquait leur plus grande mobilité, l'endurance et la tolérance dans une gamme de conditions. Cependant, cette stratégie est énergétiquement coûteuse et n'a aucune trace virtuelle dans les archives fossiles, par conséquent, le rythme et le mode d'endothermie restent controversés au cours de l'évolution des vertébrés.
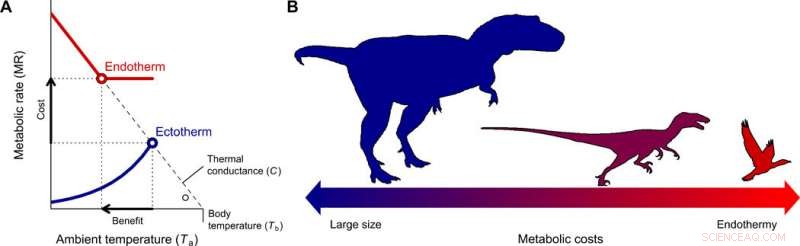
L'évolution de l'endothermie et de la miniaturisation dans la lignée des théropodes conduisant aux oiseaux. (A) Le coût-bénéfice du passage de l'ectothermie à l'endothermie pour différentes gammes de taille corporelle a été quantifié avec le modèle Scholander-Irving, qui décrit comment une élévation du métabolisme au repos (coût) augmente la niche thermique Tb − Ta (bénéfice). Parce qu'il n'y a pas de gradient thermique entre l'organisme et l'environnement en l'absence de production de chaleur, cette courbe coupe l'abscisse à Tb =Ta lorsque MR =0 (8). Les lignes bleues et rouges continues représentent les courbes métaboliques d'un ectotherme et d'un endotherme typiques, respectivement, et les symboles vides représentent le gradient thermique maximal Tb − Ta possible avec des taux métaboliques au repos, utilisé dans notre modèle (Eq. 2). (B) Une réduction de la taille du corps, conforme à celui décrit des théropodes ancestraux aux oiseaux basaux (22), constitue le chemin évolutif de moindre résistance car les coûts énergétiques d'être grand sont échangés contre ceux d'être endothermique. Crédit :Avancées scientifiques, doi:10.1126/sciadv.aaw4486
Pour comprendre l'origine et la raison de l'endothermie au cours de l'évolution des oiseaux et des mammifères, les scientifiques se sont penchés sur deux questions fondamentales. Quels sont les coûts et bénéfices de cette stratégie par rapport à l'ectothermie ? Et quelles conditions ont favorisé une transition vers l'endothermie ? Rezende et al. a répondu aux questions en utilisant le modèle Scholander-Irving de transfert de chaleur, utilisé pour étudier la thermorégulation dans les endothermes depuis plus de 60 ans. La relation était rarement utilisée pour les ectothermes, mais puisque tous les organismes vivants produisent de la chaleur endogène, le modèle est resté applicable en régime permanent thermique. Ils ont fait cette hypothèse cruciale pour éviter d'utiliser des modèles dynamiques complexes qui sont souvent appliqués à l'ectothermie, ce qui pourrait rendre les analyses actuelles difficiles.
Ils ont quantifié les coûts de l'endothermie en tant que dépense énergétique indépendante de la masse, où les avantages comprenaient une plus grande mobilité et une plus grande efficacité de recherche de nourriture, évitement des prédateurs, la tolérance et la colonisation d'un large éventail de conditions environnementales, avec des taux de croissance et une homéostasie accrus. Rezende et al. quantifié la niche thermique que les organismes pourraient occuper et son expansion pour estimer le bénéfice net de l'endothermie. Ils ont ensuite calculé le rapport coût-bénéfice de l'adoption d'un mode de vie endothermique en référence à l'ancêtre ectotherme et au descendant endothermique. Comme proposé à l'origine par le biologiste évolutionniste Bran K. McNab, lorsque les scientifiques ont reproduit ces calculs avec des estimations exactes de la taille du corps, les résultats ont montré que des tailles plus petites réduisaient les coûts énergétiques pour évoluer vers l'endothermie.
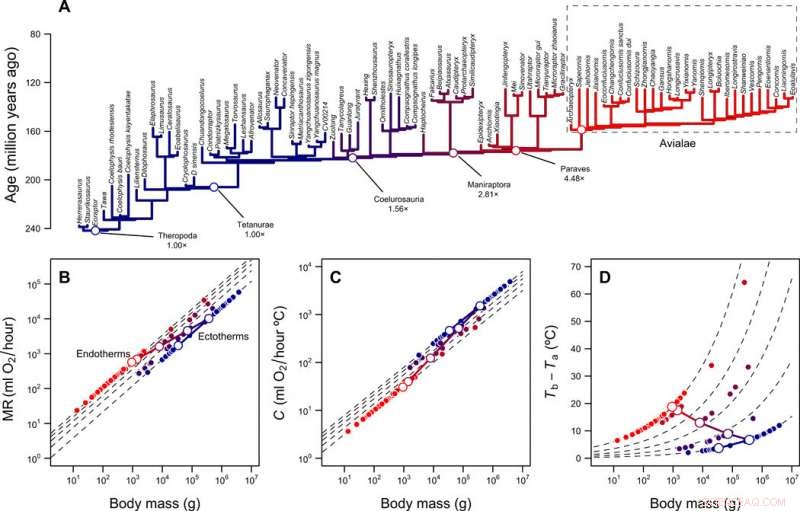
Reconstruction des niveaux métaboliques et niche thermique des théropodes. (A) phylogénie des théropodes avec des branches codées par couleur selon les niveaux métaboliques reconstruits. (B) Échelle du taux métabolique par rapport à la masse corporelle pour les ectothermes (MR =0,68 masse0,75) et les endothermes (MR =3,4 masse0,75) et la trajectoire prévue de la lignée de tiges d'oiseaux pendant la transition de l'ectothermie à l'endothermie. Les lignes pointillées montrent les différences de pli entre les ectothermes et les endothermes (1× à 5×); les symboles ouverts et fermés représentent les valeurs reconstruites pour la lignée des tiges d'oiseaux et les extrémités de la phylogénie, respectivement. (C) Échelle de la conductance thermique C et de la masse corporelle pour les ectothermes (C =2,5 masse0,5) et les endothermes (C =1,0 masse0,5), plier les différences de 2,5× à 1×. (D) gradient thermique et différences de plis calculés avec l'équation. 1 (dans la publication) et les valeurs en (B) et (C). Les trajectoires log-log linéaires reliant MR et C de l'ancêtre ectotherme et du descendant endothermique, ainsi que la trajectoire résultante en gradient thermique, sont représentés par des lignes continues. Crédit :Avancées scientifiques, doi:10.1126/sciadv.aaw4486
L'équipe a exploré comment ce modèle de transfert de chaleur s'est combiné avec des phylogénies et des reconstructions de la taille du corps, éclairer l'évolution de l'endothermie chez les oiseaux et leurs ancêtres théropodes. Rezende et al. estimé les coûts de l'évolution de l'endothermie le long de la lignée de tiges d'oiseaux en utilisant des tailles corporelles ancestrales reconstruites, sur la base des archives fossiles. Pour quantifier les coûts énergétiques dans des scénarios alternatifs, ils ont simulé l'évolution de la taille corporelle le long de la lignée et obtenu la distribution du coût par degré dans le modèle. Ils ont supposé un modèle d'Ornstein-Uhlenbeck (OU) non orienté entre une masse de 10 g et 100, 000 kg avec un taux d'évolution moyen équivalent à ceux rapportés pour les théropodes. Les simulations ont indiqué une diminution marquée des coûts énergétiques par degré avec la miniaturisation. Les scientifiques ont expliqué les coûts réduits en utilisant deux phénomènes.
D'abord, l'expansion de la niche thermique basée sur une augmentation du taux métabolique (MR) était disproportionnellement plus élevée chez les ectothermes plus grands en raison de leur capacité à maintenir une température corporelle élevée (T
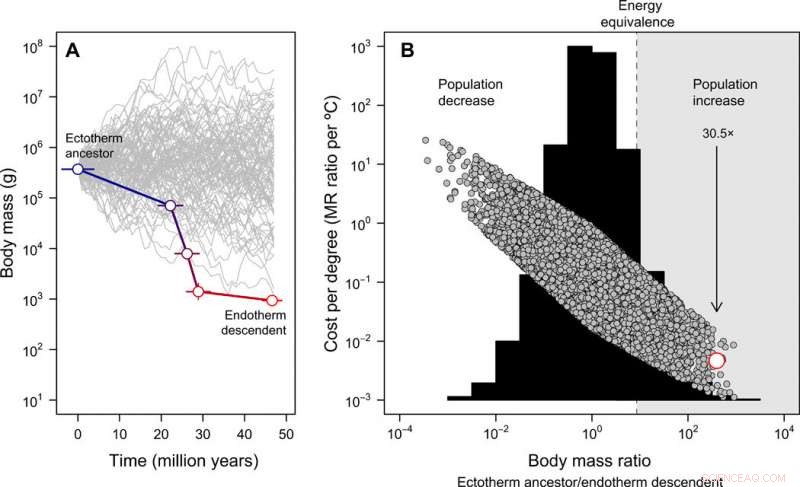
Évolution de la taille corporelle et coût-bénéfice de l'endothermie. (A) La miniaturisation des tétanures aux oiseaux basaux déduite des archives fossiles, contraste avec 100 trajectoires de taille simulées à partir de la même taille de corps ancestrale à des fins d'illustration (notez que pour le modèle nul complet suivant, la taille du corps ancestral peut varier). L'erreur représente l'écart-type dans les valeurs reconstruites sur 20 arbres candidats. (B) La distribution de fréquence des rapports de masse corporelle obtenus sur 10, 000 trajectoires de taille corporelle simulées (histogramme) et les coûts énergétiques pour faire évoluer l'endothermie exprimés par degré Celsius (Eq. 2 dans la publication) sous ce modèle nul (symboles gris). Dans ce cas, la taille du corps ancestrale a été obtenue à partir d'une distribution uniforme comprise entre 10 g et 100, 000kg. L'estimation empirique de la lignée de tiges d'oiseaux est indiquée en rouge. La région dans laquelle une réduction de la taille du corps compenserait les coûts énergétiques de l'évolution de l'endothermie, permettre à la population d'augmenter dans un scénario de ressources constantes, est surligné en gris. La flèche représente l'augmentation attendue de la population, étant donné la réduction de la taille corporelle observée dans la lignée de tiges d'oiseaux à mesure que l'endothermie évoluait. Ces analyses indiquent que les coûts énergétiques pour faire évoluer l'endothermie sont réduits avec la miniaturisation et, par conséquent, la taille de la population peut avoir augmenté malgré les coûts métaboliques d'un mode de vie endothermique. Crédit :Avancées scientifiques, doi:10.1126/sciadv.aaw4486
La réduction de la taille de la lignée de tiges d'oiseaux, correspondait étroitement au chemin théorique de moindre résistance pour l'évolution de l'endothermie, puis Rezende et al. reconstruit comment le phénomène aurait pu se dérouler dans la phylogénie des thérapodes. Le processus a indiqué l'augmentation du MR (taux métabolique) couvrant la majeure partie du Jurassique inférieur et moyen (il y a environ 180 à 170 millions d'années), impliquant des groupes de thérapodes où la présence de protoplumes et de plumes était déjà omniprésente. Ils ont également suggéré des taux métaboliques très divers à travers les lignées concurrentes de Coelurosauria, Maniraptora et Paraves, ce qui pourrait expliquer l'émergence et la diversification de ces groupes au Jurassique supérieur. Les travaux pourraient également expliquer la diversité anormalement élevée des Coelurosauria à des tailles corporelles intermédiaires (30 à 300 kg) par rapport aux autres groupes de dinosaures. L'équipe de recherche a en outre démontré un modèle de rayonnement adaptatif remplissant une niche chez les dinosaures du Mésozoïque avec des taux exceptionnels de réduction de la taille du corps dans la lignée des tiges des oiseaux (c'est-à-dire dans les nœuds basaux de Coelurosauria et Paraves), cependant, ils n'ont pas proposé d'hypothèse évolutionniste appropriée pour expliquer le résultat.
De cette façon, Enrico L. Rezende et ses collègues ont observé deux phénomènes exceptionnels au cours de l'évolution des oiseaux; (1) une miniaturisation soutenue (mais pas nécessairement progressive) s'étalant sur des millions d'années et (2) l'émergence de l'endothermie. Ils ont utilisé des reconstructions pour suggérer une évolution concomitante de l'endothermie avec une diminution de la taille le long de la lignée de la tige de l'oiseau, comme déjà proposé pour les mammifères. Les clades thérapodes correspondants présentaient un spectre complet de RM. Les interprétations indiquent que l'endothermie a précédé l'évolution du vol et que la réduction marquée de la taille du corps au cours de l'évolution de la lignée des tiges des oiseaux s'est accompagnée d'un changement majeur des niveaux métaboliques.
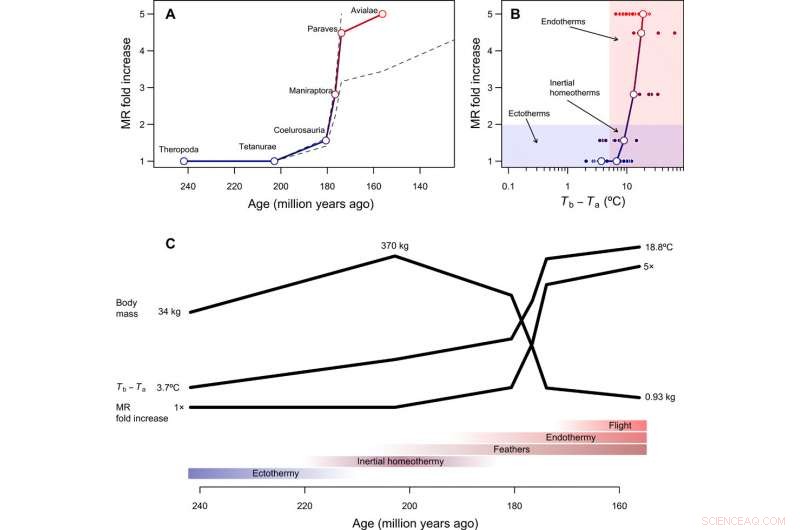
Tempo et mode dans l'évolution de l'endothermie. (A) Cours temporel reconstruit de l'évolution métabolique dans la lignée de tiges d'oiseaux, avec des lignes pointillées montrant comment les reconstructions changent en supposant que Paraves ou Neornithes étaient entièrement endothermiques au lieu de l'oiseau basal [pour les calculs avec Neornithes, nous avons supposé une taille corporelle de 150 g sur la base d'estimations pour Vegavis et d'une estimation temporelle d'il y a 100 Ma]. Le facteur d'augmentation du MR a été calculé en divisant le MR reconstruit pendant la transition vers l'endothermie par le MR attendu pour un ectotherme de taille similaire et est donc sans dimension et indépendant de la taille du corps. (B) Le chemin évolutif de moindre résistance de l'ectothermie à l'endothermie comprend l'homéothermie inertielle comme étape de transition, suivi d'une augmentation du métabolisme concomitante à une réduction de la taille. (C) Séquence hypothétique de transitions évolutives dans la lignée des tiges d'oiseaux, qui combine les résultats de cette étude avec des reconstructions phylogénétiques des structures épidermiques et de la capacité de vol actif (voir le texte principal). Crédit :Avancées scientifiques, doi:10.1126/sciadv.aaw4486
Le scénario proposé expliquait pourquoi les théropodes privilégiaient systématiquement un mode de vie coûteux (taux de renouvellement énergétique plus élevé) malgré ses coûts énergétiques, et une miniaturisation soutenue précédant l'origine des oiseaux. Par ailleurs, un état d'homéothermie inertielle constituait un état de transition nécessaire avec de faibles coûts métaboliques. Si les grands théropodes ancestraux étaient physiologiquement attachés à l'homéothermie, Rezende et al. alors attendez-vous à ce qu'ils soient sortis de l'allométrie métabolique ectothermique (relation de la taille du corps à la forme, anatomie, physiologie et comportement) avec miniaturisation. Au cours de l'évolution, il est raisonnable de s'attendre à ce que les lignées exploitent des niches nouvellement ouvertes et finissent par se diversifier. Les résultats de ce travail sont préliminaires; ils soulignent simplement qu'il y a probablement plus à l'évolution de l'endothermie chez les archosaures, dinosaures et oiseaux d'aujourd'hui. Les résultats forment une hypothèse de travail qui peut être combinée dans des études futures avec des analyses antérieures sur l'évolution de la taille et d'autres caractéristiques dans la lignée des tiges d'oiseaux pour former des interprétations bien définies des séquences temporelles au cours des transitions évolutives clés.
© 2020 Réseau Science X