La fonction principale de tous les organismes vivants, depuis la position impartiale de la survie des espèces, est de propager avec succès du matériel génétique aux générations suivantes. Une partie de cette tâche, bien sûr, reste en vie et en bonne santé assez longtemps pour s'accoupler et se reproduire. En raison de ces réalités, les unités fondamentales des êtres vivants, les cellules, ont deux fonctions principales: faire des copies identiques d'elles-mêmes pour maintenir la croissance, effectuer des réparations et prendre en charge d'autres fonctions quotidiennes au niveau des tissus, des organes et de l'ensemble organisme; et la génération de cellules spécialisées appelées gamètes qui se combinent avec les gamètes d'autres organismes de l'espèce pour générer une progéniture.
Le processus de réplication de cellules entières pour produire des cellules filles identiques comme la mitose, et il se produit dans tous les eucaryotes, qui sont les animaux, les plantes et les champignons (les procaryotes, qui sont presque tous des bactéries, se reproduisent par fission binaire, semblable à la mitose mais plus simple). La génération de gamètes se produit uniquement dans les gonades et est appelée méiose. La mitose et la méiose sont subdivisées en cinq phases, qui dans le cas de la méiose comprennent deux tours de chaque phase par cellule d'origine, car la méiose entraîne quatre nouvelles cellules plutôt que deux. La première et la plus longue de ces phases est appelée prophase, qui, dans la méiose I, est divisée en cinq phases.
Qu'est-ce que le "matériel génétique"?
Tous les êtres vivants sur Terre ont de l'ADN, ou l'acide désoxyribonucléique, comme matériel génétique. L'ADN fait partie d'une paire d'acides nucléiques qui existent dans les systèmes vivants, l'autre étant l'acide ribonucléique (ARN). Ces deux macromolécules - ainsi nommées parce qu'elles sont constituées d'un grand nombre d'atomes, dans ce cas disposés en longues chaînes de sous-unités répétitives appelées nucléotides - sont absolument critiques, bien que de manières différentes. L'ADN, porteur au niveau racine de l'information génétique, est nécessaire pour produire de l'ARN, mais l'ARN se présente sous diverses formes et est sans doute plus polyvalent.
Les sous-unités à partir desquelles l'ADN et l'ARN sont fabriqués sont appelées nucléotides . Chacun d'eux se compose de trois parties: un sucre à cinq atomes de carbone qui comprend une structure cyclique centrale pentagonale (dans l'ADN, ce sucre est le désoxyribose; dans l'ARN, c'est le ribose, qui a un atome d'oxygène supplémentaire), un groupe phosphate et un azoté ( riche en atomes d’azote). Chaque nucléotide a seulement une telle base, mais ils viennent en quatre saveurs pour chaque acide nucléique. L'ADN comprend l'adénine (A), la cytosine (C), la guanine (G) et la thymine (T); L'ARN comprend les trois premiers mais remplace l'uracile (U) par la thymine. Étant donné que toute la variation entre les nucléotides est due à des différences dans ces bases et que les acides nucléiques sont constitués de longues chaînes de nucléotides, toute la variation entre les brins d'ADN et entre l'ADN dans différents organismes est due à la variation de ces bases. Ainsi, les brins d'ADN sont écrits en termes de leurs séquences de base, comme AAATCGATG.
L'ADN existe dans les cellules vivantes sous la forme d'une hélice double brin, ou en forme de tire-bouchon. Ces brins sont liés par des liaisons hydrogène entre par leurs bases azotées à chaque nucléotide; Un appariement unique avec T et C un appariement unique avec G, donc si vous connaissez la séquence d'un brin, vous pouvez facilement prédire la séquence de l'autre, appelée brin complémentaire.
Lorsque l'ARN messager (ARNm) est synthétisé à partir d'ADN dans un processus appelé transcription, l'ARNm produit est complémentaire du brin d'ADN matrice, et est donc identique au brin d'ADN non utilisé comme matrice, sauf pour U apparaissant dans l'ARNm où T apparaît dans l'ADN. Cet ARNm se déplace du noyau des cellules où il est fabriqué vers le cytoplasme, où il "trouve" des structures appelées ribosomes, qui fabriquent des protéines en utilisant les instructions de l'ARNm. Chaque séquence de trois bases (par exemple, AAU, CGC), appelée codon triplet, correspond à l'un des 20 acides aminés, et les acides aminés sont les sous-unités de protéines entières de la même manière que les nucléotides sont les sous-unités des acides nucléiques.
Organisation de l'ADN dans les cellules
L'ADN lui-même apparaît rarement dans les êtres vivants par lui-même. La raison en est simplement la quantité phénoménale nécessaire pour porter les codes de toutes les protéines qu'un organisme doit fabriquer. Une seule copie complète de votre propre ADN, par exemple, aurait une longueur de 6 pieds si elle était étirée de bout en bout, et vous avez une copie complète de cet ADN dans presque toutes les cellules de votre corps. Étant donné que les cellules ne font que 1 ou 2 microns (millionièmes de mètre) de diamètre, le niveau de compression nécessaire pour emballer votre matériel génétique dans un noyau cellulaire est astronomique.
La façon dont votre corps le fait est en cloutant votre ADN avec des complexes protéiques appelés octamères d'histone pour créer une substance appelée chromatine, qui est environ deux tiers de protéines et un tiers d'ADN. Bien que l'ajout de masse pour réduire la taille semble contre-intuitif, pensez-y à peu près de la même manière qu'un grand magasin payant des agents de sécurité pour éviter la perte d'argent par le vol à l'étalage. Sans ces histones relativement lourdes, qui permettent un pliage et un bobinage très étendus de l'ADN autour de leurs noyaux, l'ADN n'aurait aucun moyen de se condenser. Les histones sont un investissement nécessaire à cette fin.
La chromatine elle-même est divisée en molécules discrètes appelées chromosomes. Les humains ont 23 chromosomes distincts, 22 d'entre eux étant numérotés et le reste étant un chromosome sexuel (X ou Y). Toutes vos cellules, à l'exception des gamètes, ont deux chromosomes numérotés et deux chromosomes sexuels, mais ceux-ci ne sont pas identiques, simplement appariés, car vous recevez l'un de chacun de votre mère et l'autre de votre père. Les chromosomes correspondants hérités de chaque source sont appelés chromosomes homologues; par exemple, vos copies maternelles et paternelles du chromosome 16 sont homologues.
Les chromosomes, dans les cellules nouvellement formées, existent brièvement sous une forme simple et linéaire avant de se répliquer en préparation de la division cellulaire. Cette réplication entraîne la création de deux chromosomes identiques appelés chromatides sœurs, qui sont liés à un point appelé centromère. Dans cet état, alors, tous les 46 de vos chromosomes ont été dupliqués, ce qui donne 92 chromatides en tout.
Aperçu de la mitose
Mitose, dans laquelle le contenu des noyaux des cellules somatiques (c.-à-d. " la division quotidienne des cellules, ou non-gamètes, comprend cinq phases: prophase, prométaphase, métaphase, anaphase et télophase. La prophase, discutée en détail sous peu, est la plus longue d'entre elles et consiste principalement en une série de déconstructions et de dissolutions. En prométaphase, les 46 chromosomes commencent à migrer vers le milieu de la cellule, où ils formeront une ligne perpendiculaire à la direction dans laquelle la cellule sera bientôt séparée. De chaque côté de cette ligne, appelée plaque de métaphase, se trouvent des structures appelées centrosomes; à partir de ces fibres de protéines rayonnantes appelées microtubules, qui forment le fuseau mitotique. Ces fibres se connectent aux centromères des chromosomes individuels de chaque côté à un point appelé le kinétochore, s'engageant dans une sorte de tiraillement de guerre pour s'assurer que les chromosomes, ou plus précisément leurs centromères, forment une ligne parfaitement droite le long de la plaque de métaphase. (Imaginez un peloton de soldats passant de rangées et de colonnes reconnaissables - une sorte de "prométaphase" - à une formation rigide et prête pour l'inspection - l'équivalent de "métaphase".)
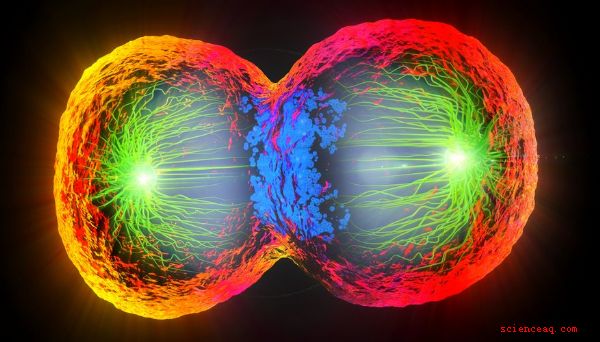
En anaphase, le phase de mitose la plus courte et la plus dramatique, les fibres du fuseau séparent les chromatides au niveau de leurs centromères, avec une chromatide attirée vers le centrosome de chaque côté. La cellule qui va bientôt se diviser semble désormais oblongue au microscope, étant "plus grosse" de chaque côté de la plaque de métaphase. Enfin, en télophase, deux noyaux filles sont complètement formés par l'apparition de membranes nucléaires; cette phase est comme la prophase inversée. Après la télophase, la cellule elle-même se divise en deux (cytokinèse).
Aperçu de la méiose
La méiose se déroule dans des cellules spécialisées des gonades (testicules chez les mâles, ovaires chez les femelles). Contrairement à la mitose, qui crée des cellules "quotidiennes" à inclure dans les tissus existants, la méiose crée des gamètes, qui fusionnent avec les gamètes du sexe opposé dans la fécondation.
La méiose est divisée en méiose I et méiose II. Dans la méiose I, au lieu que les 46 chromosomes forment une ligne le long de la plaque de métaphase comme dans la mitose, les chromosomes homologues se "traquent" et s'associent, échangeant de l'ADN dans le processus. Autrement dit, le chromosome maternel 1 est lié au chromosome paternel 1 et ainsi de suite pour les 22 autres chromosomes. Ces paires sont appelées bivalents.
Pour chaque bivalent, le chromosome homologue du père vient se poser d'un côté de la plaque métaphase, et le chromosome homologue de la mère repose de l'autre. Cela se produit indépendamment dans chaque bivalent, donc un nombre aléatoire de chromosomes d'origine paternelle et maternelle s'enroule de chaque côté de la plaque de métaphase. Les processus d'échange d'ADN (alias recombinaison) et d'alignement aléatoire (alias assortiment indépendant) assurent la diversité de la progéniture en raison de la gamme pratiquement illimitée d'ADN qui se traduit par la formation de gamètes.
Lorsque la cellule subissant la méiose I se divise , chaque cellule fille possède une copie répliquée des 23 chromosomes, plutôt que 46 chromatides à la mitose. Les 46 centromères ne sont donc pas perturbés au début de la méiose II.
La méiose II est, à toutes fins pratiques, une division mitotique, car les chromatides de la méiose I se séparent aux centromères. Le résultat final des deux stades de la méiose est de quatre cellules filles en deux paires identiques différentes, chacune avec 23 chromosomes simples. Cela permet la conservation de 46 chromosomes lorsque les gamètes mâles (spermatocytes) et les gamètes femelles (ooctyes) se joignent à la fécondation.
Prophase dans la mitose
La prophase occupe plus de la moitié de la mitose. La membrane nucléaire se décompose et forme de petites vésicules, et le nucléole à l'intérieur du noyau se désintègre. Le centrosome se divise en deux, les composants résultants s'installant sur les côtés opposés de la cellule. Ces centrosomes commencent alors à générer des microtubules qui se déploient vers la plaque de métaphase, semblable, peut-être, à la façon dont une araignée génère sa toile. Les chromosomes individuels deviennent entièrement compacts, ce qui les rend plus reconnaissables au microscope et permet une visualisation facile des chromatides sœurs et du centromère entre elles.
Prophase dans la méiose
La prophase de la méiose I comprend cinq étapes. Dans la phase leptotène, toutes les structures des chromosomes homologues non encore appariés commencent à se condenser, semblable à ce qui se produit en prophase dans la mitose. En phase zygotène, les chromosomes homologues s'associent dans un processus appelé synapsis, avec une structure appelée complexe synaptonémal se formant entre les homologues. Dans la phase pachytène, la recombinaison entre les chromosomes homologues se produit (également appelée "croisement"); Pensez à cela lorsque vous échangez peut-être une chaussette et un chapeau avec un frère ou une sœur à qui vous ressemblez étroitement en apparence et en robe. Dans la phase diplotène, le bivalent commence à se séparer, mais les homologues restent physiquement liés au niveau de leurs chiasmes. Enfin, en diakinésie, les chromosomes continuent de s'éloigner, les chiasmes se déplaçant vers leurs extrémités.
Il est essentiel de reconnaître que sans méiose, et sans les événements de la prophase I en particulier, très peu de variation entre les différents les organismes seraient évidents. Le brassage du matériel génétique qui se produit dans cette phase est l'essence même de la reproduction sexuelle.
La prophase II, qui se produit dans les cellules filles non identiques formées par la méiose I, voit à nouveau les chromosomes individuels se condenser en des formes reconnaissables , la membrane nucléaire se dissolvant à mesure que se forme le fuseau mitotique.